
Qualitative quantitative models
A systems view of calcium homeostasis and its implications

Of the many examples of homeostatic systems in Cannon’s Wisdom of the Body, one of the cleanest and most beautiful examples is the regulation of Calcium. In 1932, it was evident that calcium facilitated bone growth, muscle control, and blood coagulation. Moreover, the concentration of calcium in the blood needed to be constant. People with low calcium blood levels would suffer from muscle stiffness and spasms. High calcium concentration would result in increased blood viscosity. By what mechanism did the body maintain the sweet spot of ideal calcium concentration?
Though there wasn’t a complete picture, the systems involved in calcium homeostasis were clear to Cannon. The parathyroid gland played a central role, as removing or damaging it caused dysregulation. Rodent experiments showed that calcium levels could be restored by surgically adding parathyroid tissue. Human studies demonstrated parathyroid extract was an effective treatment of hypocalcemia.1 The bones were also involved, as they held larger stores of calcium when animals were fed high calcium diets. Vitamin D was known to help facilitate absorption of calcium from the intestines, and vitamin D supplementation had effects similar to those of parathyroid extract.
How do these systems work in tandem? The first clean systems-level description I’ve found didn’t appear until 2002. In the Journal of Theoretical Biology, Hana El-Samad, Jesse Goff, and Mustafa Khammash derived a relatively simple physiological model I’ll describe here. Their model highlights a key feature of homeostatic systems.
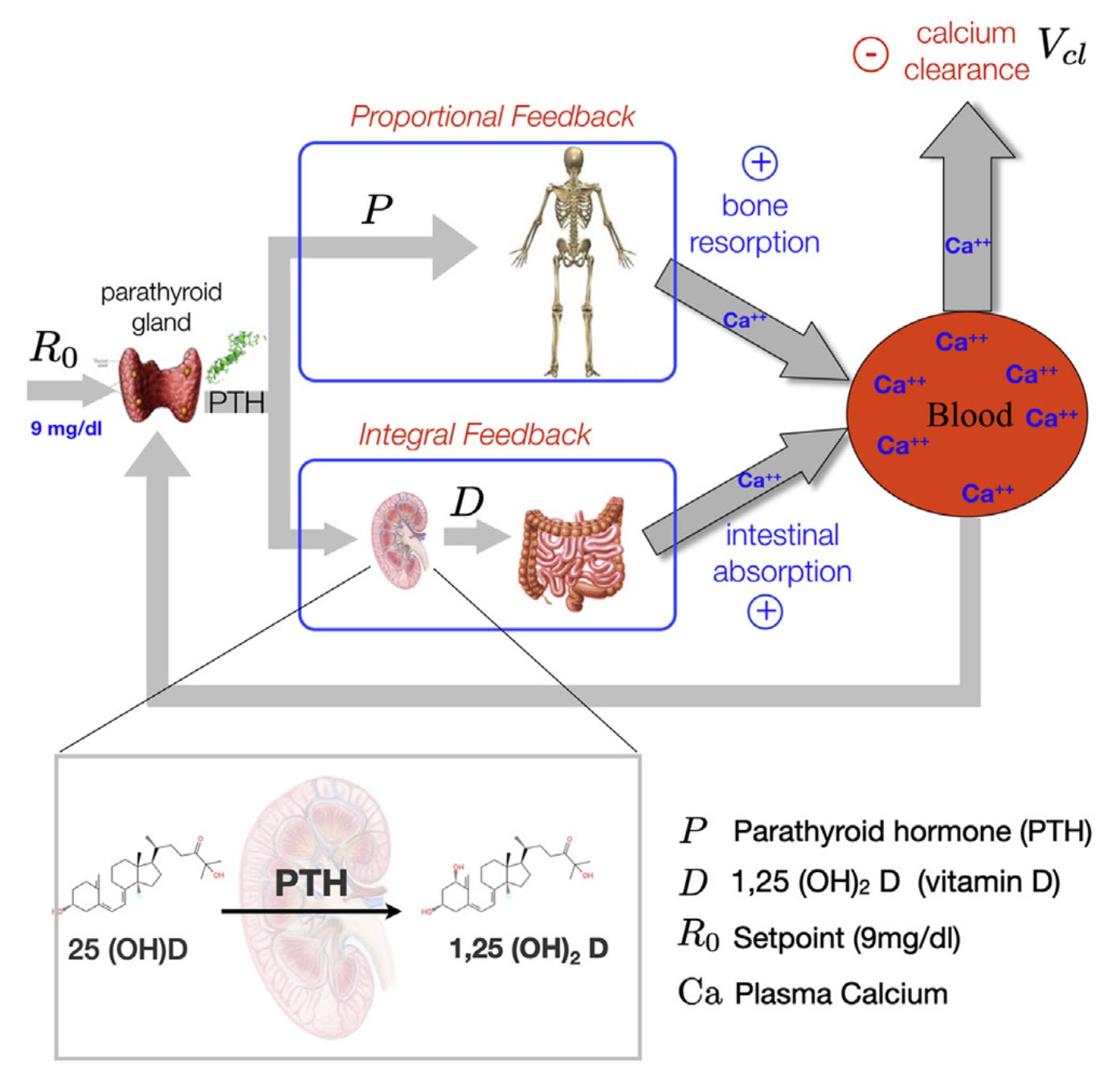
The systems involved in the model are precisely those identified by Cannon. The calcium level in the bloodstream changes based on three signals. First, an external demand can remove calcium. Second, calcium can move from the bones to the blood. Third, calcium can move from the intestine to the blood.
The amount of calcium extracted from the bones is proportional to the level of parathyroid hormone (PTH) in the blood. The amount extracted from the intestines is proportional to the level of active vitamin D in the blood. And the amount of active vitamin D added to the blood at any time is proportional to the amount of PTH. PTH stimulates the kidney to activate vitamin D.
Now, we close the loop: the source of the PTH in the blood is the parathyroid gland. Experimental evidence shows the amount of PTH in the blood scales linearly with the deviation of the calcium concentration from its homeostatic setpoint. That is, the PTH in the bloodstream serves as an error signal.
Put this picture together: some external demand depletes calcium in the blood. In response, the parathyroid gland releases PTH. The rate of calcium resorption from the bone then increases linearly, and the rate of the rate of calcium extraction from the intestines increases linearly. This “double rate” system is the critical one. At steady state, the body must add a constant rate of calcium to the blood. For this rate to be constant, the change of the rate has to be zero. This second-order effect, governed by the vitamin D production of the kidneys, is directly proportional to the setpoint error in the calcium level. Thus, we must have that the setpoint error in calcium concentration is driven to a constant level.
The cascading effect mitigated by the kidney was critical. If only the bones were involved, we would know that the steady-state calcium level would be a constant, but we wouldn’t know which constant. With the two-stage system, a constant calcium level implies that the value equals the setpoint.
There’s another way to see that the system has to converge to a state where the setpoint error is zero. The amount of activated vitamin D in the blood increases whenever the setpoint error is not zero. The only way for that active vitamin D level to not increase forever is for the set point error to go to zero.
The two-stage structure with the kidney results in the rate of intestinal calcium extraction being proportional to the integral of the PTH level over time. In fact, the vitamin D level is the integral of the setpoint error over time. Hence, the change in the calcium level in response to external demand is equal to a linear combination of the setpoint error signal and its integral. In control theory, we call such a regulator a proportional-integral controller.
This picture, from Khammash’s 2021 survey on perfect adaptation, illustrates the feedback loop.
I purposefully wrote this blog without equations because, in some sense, they don’t matter. A high-level systems view illustrates how calcium homeostasis is maintained. A network of biochemical reactions regulates the calcium level, and we can view the regulatory cascade as a proportional-integral controller. This is a qualitative quantitative result. We are making a few assumptions about linearity, but these are fairly mild and consistent with experiments.
Moreover, the model’s equations are underspecified. They depend on at least four parameters whose values I don’t know. The remarkable fact is that no matter what the values of these parameters are, the system must converge to the correct setpoint if it converges at all. If there’s a setpoint, integral control finds it. If calcium demand unexpectedly changes, the body adapts, recovering the same calcium concentration as before the shock. The steady-state values of calcium storage in bones and activated vitamin D levels in the bloodstream change in the new environment to ensure the calcium concentration does not. This adaptation to unknown futures is a remarkable property of integral control, and I’ll discuss it in the next post.
I love medical terminology where you just concatenate prefixes and suffixes together to make it sound like you understand something. Low calcium = hypocalcemia.
I wonder if we are confusing the broader community by calling this integral control. The essence is that the steady equations are structured such that they imply regulated variable = reference. This may be because there is a specific variable whose derivative is equal to reference minus regulated variable (an explicit integrator) or you can do a change of variables to reach that specific form, but the general implication I wrote is much more general and easy enough to explain to a biologist with a couple of examples. Why do we need to see an integrator explicitly, other than making a connection to control theory?
Elegant representation of elegant biology. As a cardiologist, a similar analysis of blood pressure or heart rate would be fun. Sneak in some calculus if you can